Access and read the PDF with inline linkages, and reference list here.
Abstract
The global warming potential of methane (CH4) is about 30 times stronger than that of carbon dioxide (CO2) over a century timescale. Methane emission is hypothesized to have contributed to global climate change events and mass extinctions during Earth’s history. Therefore, the study of CH4 production processes is critically important to the understanding of global climate change.
It has been a dogma that biogenic CH4 detectable in the oceans originates exclusively from the anaerobic metabolic activity of methanogenic archaea in hypoxic and anoxic environments, despite reports that many oxic surface and near-surface waters of the world’s oceans are CH4-supersaturated, thereby rendering net sea-to-air emissions of CH4.
The phenomenon of CH4 production in oxic marine waters is referred to as the “ocean methane paradox”. Although still not totally resolved, recent studies have generated several hypotheses regarding the sources of CH4 production in oxic seawater.
This review will summarize our current understanding of the importance of CH4 in the global climate and analyze the biological processes and their underpinning mechanisms that lead to the production of CH4 in oxic seawater environments. We will also tentatively explore the relationships of these microbial metabolic processes with global changes in climate and environment.
Introduction
Global warming, caused mainly by the rapid increase of anthropogenic greenhouse gas emissions, is dramatically changing the Earth’s climate and environment. In May 2013, for the first time in human history, the daily average atmospheric carbon dioxide (CO2) concentration exceeded 400 ppm (parts per million), as measured at the Mauna Loa Observatory, Hawaii (Showstack, 2013). This atmospheric CO2 concentration is likely to be the highest during the last 3.4 Myr, 15 Myr, or even 34 Myr of the Earth’s history (Beerling and Royer, 2011; Bijma et al., 2013; BrighamGrette et al., 2013).
The current anthropogenic carbon release rate (~10 Pg C yr−1) is nearly 10 times larger than that of the Palaeocene-Eocene Thermal Maximum (PETM), which presumably had the highest natural carbon release rate (~1.1 Pg C yr−1) during the past 66 Myr on Earth (Zeebe et al., 2016). Excessive anthropogenic CO2 production and emission, mainly due to fossil fuel combustion, deforestation, and cement production, has been identified as the main cause of global climate change (Bijma et al., 2013). There have been a huge number of studies of CO2-related climate change and its environmental impacts. However, in addition to CO2, other greenhouse gases such as methane (CH4) and nitrous oxide (N2O) also contribute to global warming (Dickinson and Cicerone, 1986; Ravishankara et al., 2009; Dlugokencky et al., 2011; Montzka et al., 2011; Carpenter et al., 2012).
Emissions of CH4 and N2O have also increased rapidly during the last few centuries, mainly due to anthropogenic activities (Forster et al., 2007; Bakker et al., 2014). Although it may be possible to keep anthropogenic CO2 emission under control in the future, the global warming effect of other greenhouse gases will still result in a continuous increase of the global temperature (Solomon et al., 2010; Montzka et al., 2011).
To achieve a successful global change mitigation it is necessary to take all of the major greenhouse gases into consideration. Methane is the second most potent greenhouse gas contributing ~20% to the present-day anthropogenic forcing of well mixed greenhouse gases relative to the pre-industrial era (Wuebbles and Hayhoe, 2002; Montzka et al., 2011; Kirschke et al., 2013).
Although the atmospheric CH4 concentration (~1.83 ppm) is much lower than that of CO2 (~400 ppm), the anthropogenic CH4 radiative forcing is about 1/4 to 1/3 that of anthropogenic CO2 (Etminan et al., 2016; Ruppel and Kessler, 2017). Due to excess anthropogenic emissions, the atmospheric CH4 concentration is increasing at ~2% per year (Rasmussen and Khalil, 1981). The current atmospheric CH4 concentration is the highest over at least the past 0.8 Ma and has increased 2.5 times since the Industrial Revolution, which is mainly attributable to anthropogenic activities (Wuebbles and Hayhoe, 2002; Loulergue et al., 2008; Dlugokencky et al., 2011).
In comparison, the atmospheric CO2 concentration has increased by less than 50% during the same period. Therefore, it has been reported that CH4 plays a prominent role in global warming (Ruppel and Kessler, 2017). This viewpoint is further substantiated by the positive feedback loop between CH4 and global warming (MacDougall and Knutti, 2016). The estimation of the global CH4 emission is about 500–600 Tg yr−1 (Conrad, 2009; Ghosh et al., 2015; Tsuruta et al., 2017). Under the impact of global warming, CH4 emissions from both natural and anthropogenic sources are expected to increase (Montzka et al., 2011; Hamdan and Wickland, 2016), which in turn will exert an even more intense influence on the rise in global temperature.
Methane also plays a very important role in modulating atmospheric chemistry. In the troposphere, CH4 is oxidized by the hydroxyl radical (OH·) and this process leads to ozone pollution (Montzka et al., 2011; Prather and Holmes, 2017). Methane oxidation contributes to the buildup of stratospheric water vapor (H2O), which is also a greenhouse gas causing global warming (Isaksen et al., 2011). In addition to creating a sink for atmospheric CH4, OH· has a major role in the destruction of some other tropospheric greenhouse gases, such as hydrofluorocarbons and hydrochlorofluorocarbons (Montzka et al., 2011).
Moreover, OH· can oxidize tropospheric sulfur dioxide (SO2), forming sulfate aerosols, which have a certain cooling effect on the global climate (Montzka et al., 2011). The consumption of OH· by CH4 may thus create another positive feedback on global warming, by increasing the lifetime of atmospheric CH4 and changing the dynamics of some other tropospheric greenhouse gases (Isaksen et al., 2011; Montzka et al., 2011).
Moreover, CH4 also contributes to the increase in the atmospheric CO2 concentration, with CO2 being the end product of CH4 oxidation (Isaksen et al., 2011). Furthermore, CH4 may exert an important radiative forcing-induced long-term (>200 years) effect on the ongoing sea-level rise through thermal expansion (Zickfeld et al., 2017), even if its anthropogenic emission can be halted and even though its atmospheric lifetime is only about 9 years (Sonnemann and Grygalashvyly, 2014).
Overall, CH4 may play an increasingly more important role in the Earth’s climate and environment systems in the future. The global warming potential (GWP) of CH4 over a 100- year period is over 20 times stronger than that of CO2. However, to keep global warming under control in the near future (i.e., a couple of decades), the GWP of CH4 over a 20- year period is more relevant to consider and is over 70 times stronger than that of CO2 (Wuebbles and Hayhoe, 2002; Karthikeyan et al., 2015).
Although the task of finding a solution in this short time is tremendously challenging and difficult, there is a need to act quickly and early to avoid the danger incurred by global change and to not step over the threshold and lose the opportunity (Hansen et al., 2007; MacCracken, 2008). It has been proposed that the current anthropogenic activity-induced global warming may be irreversible and there are serious concerns that a domino effect may be triggered, causing the Earth to spiral out of balance and generating uninhabitable conditions, if we cannot act quickly and decisively to stop climate change in the near future (e.g., Frondel et al., 2002; Solomon et al., 2009; Hansen et al., 2013; Schleuning et al., 2016).
Because the steady-state lifetime of CH4 in the atmosphere is much shorter than that of CO2, the reduction of CH4 emissions (and other greenhouse gases with short lifetimes) should effectively and quickly help mitigate climate change (Montzka et al., 2011; Karol’ et al., 2013). The reduction in radiative forcing through the removal of atmospheric CH4 and other short-lived greenhouse gases is much faster than that of CO2 (Zickfeld et al., 2017).
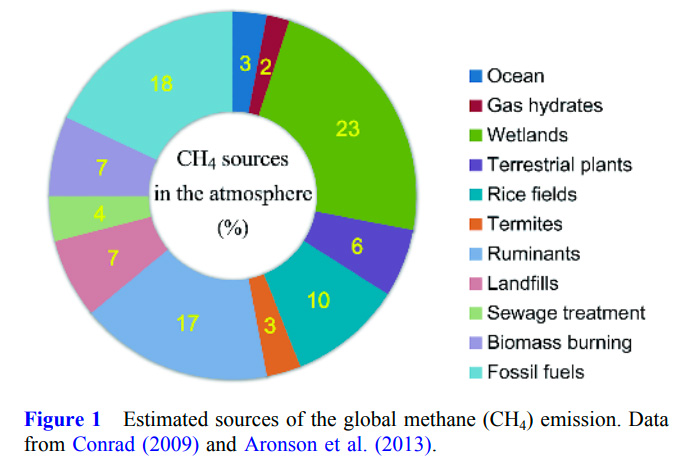
Although a diverse range of natural and anthropogenic sources of CH4 emissions have been identified (Figure 1), the mechanisms and contributions of some of the sources to atmospheric CH4 have not been clearly resolved, and this is particularly true for the ocean (Kirschke et al., 2013; Hamdan and Wickland, 2016). This situation hinders a mechanistic and comprehensive prediction of the global warming caused by CH4 and its application in climate change mitigation.
The ocean as a net source of CH4 to the atmosphere
In general, the ocean contributes to net sequestration of anthropogenic CO2. It has been estimated that ~40% of the anthropogenic CO2 emissions since the Industrial Revolution have been absorbed by the ocean (Sabine et al., 2004; McKinley et al., 2016; DeVries et al., 2017). However, the ocean is generally regarded to be a minor contributor to atmospheric CH4, based on various estimations (Bange et al., 1994; Matthews, 1994; Krüger et al., 2005; Carpenter et al., 2012).
The ocean-to-atmosphere emission rate of CH4 is estimated to be ~1.3 to 13 Tg yr−1 (Ruppel and Kessler, 2017). However, the wide range of estimations indicates that these estimations contain much uncertainty due to a lack of sufficient sampling coverage and long time-series measurements of the global ocean, and they are unable to capture the temporospatial dynamics of CH4 emissions in the ocean (Bange, 2006; Fischer et al., 2013; Hamdan and Wickland, 2016).
The sea-to-air CH4 flux varies substantially among different marine environments and the mechanisms that control the huge temporal and spatial variation of CH4 emissions are still not fully resolved (Bates et al., 1996; Ortiz-Llorente and Alvarez-Cobelas, 2012; Wilson et al., 2017). The lack of long-term measurements with sufficient temporal and spatial frequencies and resolutions, in particular at regional and global scales, hinders global marine CH4 data integration, and thus an accurate estimation of marine CH4 emissions from the global ocean. Compared to the ocean, the global wetlands are a much larger source of CH4 emissions (Bridgham et al., 2013).
A recent study reported that the average CH4 emission from wetlands on a global scale was 177.2±49.7 Tg CH4 yr−1 for the period from 2000 to 2007 (Zhang et al., 2017). As the land-ocean interface, coastal wetlands also exert a strong influence on CH4 emissions from coastal marine waters. Due to the large terrestrial, fluvial, and anthropogenic inputs of organic matter and nutrients, intense microbial activities, including methanogenesis, occur in coastal wetlands (Vizza et al., 2017; Xiao L et al., 2017).
Although coastal wetlands, defined as areas of marine water with a water depth not exceeding 6 m at low tide (Navid, 1989), account for only a small fraction of the global ocean system, their CH4 production may constitute a major source of CH4 emissions in adjacent estuarine and coastal seas (Wang et al., 2016). In the ocean, many estuarine and coastal waters are hotspots of CH4 emission. This is usually caused by intense terrestrial and sediment inputs and in situ microbial methanogenesis under hypoxic and anoxic conditions (Zhang et al., 2004, 2008; Bange, 2006; Zhou et al., 2009; Borges et al., 2016; Upstill-Goddard and Barnes, 2016; Farías et al., 2017; Sela-Adler et al., 2017; Tseng et al., 2017).
In addition to the direct transport of terrigenous CH4, river discharges usually contain high concentrations of organic matter and nutrients. This can stimulate various forms of biological production and respiration in the estuarine and coastal microbial communities and create locally and/or seasonally hypoxic and even anoxic conditions (Naqvi et al., 2010; Dang and Jiao, 2014). Estuarine and coastal seawater commonly contains high concentrations of suspended particles, likely harboring hypoxic and anoxic microenvironments inside the particles for anaerobic microbial processes (Brooks et al., 1981; Wright et al., 2012; Dang and Jiao, 2014; Dang and Lovell, 2016).
The oxygen-depleted environments and microenvironments in seawater facilitates CH4 production by anaerobic archaeal methanogens and sea-to-air CH4 emission (Offre et al., 2013; Wen et al., 2017). Although the estuarine and coastal seas only occupy a small fraction of the global ocean, they may contribute ~75% of the total marine CH4 emissions to the atmosphere (Bange et al., 1994; Reeburgh, 2007). Coastal upwelling can bring CH4 from the bottom water into surface water and contributes directly to CH4 emission (Bange et al., 1998; Capelle and Tortell, 2016; Chronopoulou et al., 2017).
In addition, the enrichment of nutrients by upwelling stimulates the biological production of organic matter and microbial community respiration, creating hypoxic and anoxic environments in seawater for anaerobic methanogenesis (Sansone et al., 2001; Wright et al., 2012; Bakun, 2017; Shepherd et al., 2017). Therefore, the hypoxic and anoxic zones of the marginal sea upwelling areas are environments of variable CH4 production and emission. Climate change may intensify coastal upwelling, particularly at high latitudes (Sydeman et al., 2014; Wang et al., 2015). This change may exert a strong impact on the structures and functions of the coastal marine ecosystems, including the intensified production and emission of greenhouse gases such as CH4 and N2O.
Sediments are a major source of seawater CH4 (Orcutt et al., 2013; Chronopoulou et al., 2017; Tseng et al., 2017; Xiao K Q et al., 2017). The anoxic conditions in bulk sediments facilitate anaerobic microbial metabolisms, including methanogenesis (Ferry and Lessner, 2008; Sela-Adler et al., 2017). Sediments in marginal seas receive high inputs of organic matter that contribute to microbiogenic CH4 production (Wen et al., 2017). Many marginal sea sedimentary environments in the world’s oceans harbor methane hydrates (also called gas hydrates).
Because they are metastable and have a high sensitivity to warming, gas hydrates are a net source of CH4 emission to the atmosphere (Conrad, 2009). Although the estimated contribution of CH4 emission (~6 Tg yr−1, ranging from 2 to 9 Tg yr−1) from marine gas hydrates to atmospheric CH4 is currently small (Ruppel and Kessler, 2017), gas hydrates have the potential to release a large amount of CH4 in a relatively short period of time, and thus they may constitute a tipping point in future global climate change (Archer et al., 2009; Carpenter et al., 2012).
Gas hydrate CH4 emission as a looming factor influencing climate change
In certain sediments, such as those harboring gas hydrates, the CH4 seepage can be phenomenal (Boetius and Wenzhöfer, 2013; Rakowski et al., 2015). For example, CH4 gas bubbles rising through seawater can reach 400 m above the seafloor where active seepage sites exist (Dang et al., 2010). Both biogenic and thermogenic CH4-producing processes can contribute to the accumulation of CH4 in marine sediments (Archer, 2007; Reeburgh, 2007; Hester and Brewer, 2009). With a favorable gas composition and under lowtemperature and high-pressure conditions, methane hydrates can form in marine sediments (Ruppel and Kessler, 2017).
It has been estimated that marine gas hydrates may contain a huge amount of CH4, ranging from 1700 to 4100000 Gt C globally (Kvenvolden, 1988). The latest estimations suggest that the inventory of marine methane hydrates may be at the lower end of the original estimates, about 1100–2000 Gt C (Archer et al., 2009; Boswell and Collett, 2011; Kretschmer et al., 2015; Ruppel and Kessler, 2017). Because the spatial distribution of sediment gas hydrates is extremely heterogeneous in the ocean and there is generally a lack of sampling coverage across the global ocean, it is not easy to make an accurate estimate of gas hydrate resources (Marín-Moreno et al., 2016), nor is it easy to estimate the CH4 emission fluxes from the globally dispersed gas hydrate sources (Matthews, 1994).
Although there may be a huge amount of CH4 dissociated from the seafloor gas hydrates every year, a variable (and usually small) fraction of hydrate CH4 may actually enter the atmosphere. It is commonly estimated that hydrate CH4 may contribute only ~2% to the pool of atmospheric CH4 (Matthews, 1994; Conrad, 2009; Hamdan and Wickland, 2016; Ruppel and Kessler, 2017). Anaerobic and aerobic methanotrophic microbes (including bacteria and archaea) consume the majority of the free CH4 pool in sediments and seawater, presumably substantially lowering the global seato-air CH4 flux (Krüger et al., 2005; Reeburgh, 2007; Knittel and Boetius, 2009; DiSpirito et al., 2016; James et al., 2016).
To facilitate CH4 metabolism, many microbial methanotrophs harbor N2-fixation genetic inventories to provide biologically fixed nitrogen (Auman et al., 2001; Dedysh et al., 2004; Pernthaler et al., 2008; Dang et al., 2009; Dekas et al., 2009; Khadem et al., 2010; Fernández-Carrera et al., 2016). This genetic and biochemical adaptation strategy indicates that marine methanotrophic microbes may have evolved in CH4-rich marine environments throughout the Earth’s history. Recent studies have identified a highly variable contribution of hydrate CH4 emissions to global warming (Biastoch et al., 2011; Kretschmer et al., 2015; Mestdagh et al., 2017).
The massive CH4 emissions putatively caused by abrupt seafloor gas hydrate dissociation are considered to be a very important factor contributing to global warming during certain historical events in the Earth’s history, including the PETM and the termination of the Marinoan “Snowball” ice age (Dickens et al., 1995, 1997; Kennett et al., 2003; Maslin et al., 2004; Svensen et al., 2004; Kennedy et al., 2008; Dickens, 2011). Catastrophic CH4 emissions associated with massive and abrupt marine gas hydrate dissociation are also hypothesized to have been involved in mass extinctions (Katz et al., 1999; Norris and Röhl, 1999; Hesselbo et al., 2000).
Although the “clathrate gun hypothesis” has been challenged (Higgins and Schrag, 2006; Sowers, 2006; Gutjahr et al., 2017), CH4 emission from dissociated gas hydrates caused by gradually warming seawater over timescales of tens of thousands of years may still have an important impact on the climate (Archer et al., 2009). Leifer et al. (2006) identified natural marine seepage blowout as a process enabling rapid climate change by sudden hydrate dissociation-related massive methane bubble releases.
Furthermore, the oxidation of CH4 released from dissociated gas hydrates consumes O2 and produces CO2, leading to ocean deoxygenation and acidification (Biastoch et al., 2011; Boudreau et al., 2015). The metastable nature of gas hydrates indicates that they are sensitive to changes in temperature and pressure, while sediment gas hydrates can be dissociated by environmental disturbances, such as a rising seawater temperature (Reagan and Moridis, 2007; Hunter et al., 2013; Ruppel and Kessler, 2017).
It has been hypothesized that CH4 emission from gas hydrate dissociation may become stronger with an increase in the global and ocean temperature (Kvenvolden, 1988). There is evidence that the ocean’s interior is getting warmer due to the impact of global warming (Masuda et al., 2010; Mora et al., 2013; Levin and Le Bris, 2015). Modeling studies have indicated that CH4 emissions from marine gas hydrates are increasing under the influence of ongoing global warming, particularly in certain sensitive environments such as the Arctic Ocean and shallow marginal seas (Reagan and Moridis, 2007; Shakhova et al., 2010; Isaksen et al., 2011; Marín-Moreno et al., 2013; Thatcher et al., 2013; James et al., 2016).
There have been observations and model predictions that CH4 is released from gas hydrates to the atmosphere in response to global warming (Archer, 2007; Thatcher et al., 2013; Stranne et al., 2017). Global change is not a simple process, and actually involves nonlinear and sometimes abrupt changes of the Earth system and its components (McNeall et al., 2011). This complexity suggests that a small change in a particular external forcing could cause large and/or irreversible changes to climate (McNeall et al., 2011).
For example, methane seepages from seafloor sediments are frequently and widely found to be episodic in many different marine environments (Stranne et al., 2017). In addition to the effects caused by the rising bottom water temperature due to global warming, there are other processes and factors in the ocean that may influence the stability and CH4 emission of sediment gas hydrates. The current climate change may eventually alter the ocean circulation pattern and the intensity of some currents.
For example, changes in the Gulf Stream intermediate water temperature could cause a rapid destabilization of gas hydrates along a broad path in the North American margin seas (Phrampus and Hornbach, 2012). Submarine volcano eruptions may release CH4 from gas hydrates if these two geological features happen to exist in the same area (Svensen et al., 2004). This is highly likely because most (~80%) of the Earth’s volcanic activity happens in submarine environments (Embley et al., 2006). Large-scale submarine landslides are a major geohazard that can trigger basin-wide tsunamis.
Earthquakes, volcanoes, hurricanes, and gas hydrate dissociation can trigger submarine slides (Masson et al., 2006; Geissler et al., 2016; Handwerger et al., 2017), which in turn may cause further large-scale gas hydrate dissociation and CH4 releases. Earthquakes can also directly cause seafloor ruptures, gas hydrate dissociation, and an intensification of methane seepage (Tsunogai et al., 2012; Fischer et al., 2013; Obzhirov, 2013; Geersen et al., 2016). Global warming may impact on some of these submarine geohazard processes.
For example, the warming of bottom seawater may destabilize gas hydrates, which in turn may lead to submarine landslides and lead to further hydrate CH4 emissions. This scenario may occur in the Arctic Ocean and some other shallow seas. In Greenland and the Antarctic, another mechanism may operate. Here, global warming has caused the shrinking of ice sheets and a rise in the affected continental slope through isostacy.
The lowered hydrostatic pressure may destabilize gas hydrates, leading to massive landslides and gas hydrate dissociation (Maslin et al., 2010). In addition, the commercial exploitation of offshore oils and gas hydrates may also trigger gas hydrate dissociation and CH4 release (Glasby, 2003; Zhang and Zhai, 2015; Fernández-Carrera et al., 2016). The positive feedback effect of hydrate CH4 emission on global warming and the various processes that could trigger large-scale “catastrophic” CH4 releases from gas hydrates has led to serious concerns about the impact of gas hydrates on the global climate (Reagan and Moridis, 2008; Archer et al., 2009; Carpenter et al., 2012; Ruppel and Kessler, 2017).
The ocean methane paradox and putative mechanisms
Many microorganisms produce greenhouse gases that exacerbate global warming. For example, it has been estimated that methanogenic archaea contribute nearly 60% of the methane emissions that are related to anthropogenic activities (Jabłoński et al., 2015). Methane was once believed to be produced exclusively by methanogenic archaea affiliated with the phylum Euryarchaeota (i.e., the Methanobacteria, Methanococci, Methanomicrobia, and Methanopyri classes) (Balch et al., 1979; Garcia et al., 2000; Liu and Whitman, 2008).
Recently, novel euryarchaeotal methanogens, such as those affiliated with the newly defined family Candidatus Methanoflorentaceae (within class Methanomicrobia), orders Methanocellales (within class Methanomicrobia) and Methanomassiliicoccales (within class Thermoplasmata), and classes Ca. Methanofastidiosa and Methanonatronarchaeia have been discovered (Sakai et al., 2008; Iino et al., 2013; Mondav et al., 2014; Nobu et al., 2016; Sorokin et al., 2017). In addition, non-euryarchaeotal methanogens, such as those affiliated with the new archaeal phyla Bathyarchaeota and Verstraetearchaeota, have also been discovered recently (Evans et al., 2015; Vanwonterghem et al., 2016).
Four distinct biochemical pathways for the production of CH4 in archaeal methanogens have been identified: (1) hydrogenotrophic methanogenesis using H2 as an electron donor to reduce CO2 for CH4 production, (2) acetoclastic methanogenesis using acetate as the substrate for CH4 disproportionation production, (3) methylotrophic methanogenesis using methylated C1 compounds (e.g., methanol, methylamines, and methylthiols) as substrates for CH4 disproportionation production, and (4) methyl-reducing methanogenesis using methylated C1 compounds as substrates and H2 and/or formate as electron donors for CH4 production by methyl group reduction (Kallistova et al., 2017). Coenzyme M (CoM) and coenzyme F420 are central cofactors in methanogenic archaea and methyl-CoM reductase is a unique key enzymatic complex that has a function in the final step of methanogenesis (Balch et al., 1979; Friedrich, 2005; Krishnakumar et al., 2008; Purwantini et al., 2014; Dziewit et al., 2015; Greening et al., 2016).
A common physiological characteristic of all the above-mentioned archaeal methanogens is that they are anaerobes, producing CH4 only in anoxic environments such as oxygen-depleted marine sediments and seawater (Valentine, 2011; Offre et al., 2013; Welte and Deppenmeier, 2014). The energy yield by traditional methanogenesis is very low (Schink, 1997; Schäfer et al., 1999). In environments with the presence of other oxidants and oxidative metabolic processes, methanogens are rarely successful competitors (Sela-Adler et al., 2017; Wen et al., 2017). Surprisingly, however, most oxic surface or nearsurface waters in the world’s oceans are supersaturated with CH4 relative to atmospheric CH4 concentrations.
This puzzling phenomenon is referred to as the “ocean methane paradox” (Reeburgh, 2007; Karl et al., 2008). There is much evidence indicating that CH4 in these waters may be produced in situ and constitutes a net flux to the atmosphere (Lamontagne et al., 1971; Scranton and Brewer, 1977). It has been estimated that the world’s open ocean CH4 emission flux to the atmosphere may be ~3.6 Tg yr−1 (Lambert and Schmidt, 1993; Bange et al., 1994), while some other studies have shown that the actual flux (~0.4 Tg yr−1) may be an order of magnitude less (Bates et al., 1996; Rhee et al., 2009).
Although the estimated contribution of the open oceans to atmospheric CH4 is highly variable among different studies, it is widely accepted that the open oceans generally constitute a net CH4 source to the atmosphere, although the underlying CH4 production mechanism remains unclear and hotly debated (Reeburgh, 2007; Karl et al., 2008; Conrad, 2009; Rakowski et al., 2015; Tseng et al., 2017).
Anoxic microenvironments associated with marine particles and zooplankton guts may host active methanogenic archaea, contributing to in situ CH4 production and accumulation in oxic bulk seawater (Oremland, 1979; Brooks et al., 1981; Cynar and Yayanos, 1991; de Angelis and Lee, 1994; Karl and Tilbrook, 1994; Tilbrook and Karl, 1995; Marty et al., 1997; Holmes et al., 2000; Reeburgh, 2007; Sasakawa et al., 2008; Ditchfield et al., 2012; Dang and Lovell, 2016). Some methanogens may have developed ecophysiological adaptations to various adverse environments. For example, some methanogens can survive oxic conditions and resume methanogenic activities when the environment is conducive (Zehnder and Wuhrmann, 1977; Jarrell, 1985; Sieburth et al., 1993; Tholen et al., 2007; Poehlein et al., 2017).
The surface waters of the ocean may experience a diel alternation of the dissolved oxygen content due to day-time phytoplankton oxygenic photosynthesis and night-time community oxygen-consuming respiration (Dang and Lovell, 2016). It may be easier to achieve hypoxic and even anoxic microenvironments in marine particles and biofilms, which would facilitate methanogenesis during the night. In addition, some environmental methanogen isolates harbor diverse adhesin-like proteins, which are useful for colonizing marine particles and other substrata (e.g., zooplankton surfaces and/or digestive tracts) (Dang and Lovell, 2016; Poehlein et al., 2017).
Recently a novel methanogen, Candidatus Methanothrix paradoxum, was identified in oxygenated soils, and it was shown to conduct acetoclastic CH4 production in the anoxic microenvironments of soil aggregates (Angle et al., 2017). The novel feature of this archaeon is its ability to compete for substrate acquisition under low acetate conditions, which provides an ecophysiological advantage over other methanogens in bulk oxic environments. Archaeal 16S rRNA gene sequences that are very similar to that of Ca. Methanothrix paradoxum are prevalent in oxic permafrost, freshwater, wetland, estuary, and marine environments (Angle et al., 2017), and some sequences were obtained from the prokaryotic communities associated with marine snow (Vojvoda et al., 2014).
The discovery of this specific methanogen indicates that the anoxic microenvironment hypothesis is likely to be valid for explaining the “ocean methane paradox”, at least for certain marine waters. Methane production also occurs in particle-free oxygenated seawater (Bogard et al., 2014), and certain studies have found that zooplankton contribute very little to oxic CH4 production (Schmale et al., 2018). The ocean methane paradox was further tentatively resolved by the finding that some marine bacteria, including cyanobacteria, can metabolize methylphosphonate (MPn), producing CH4 as a byproduct for phosphorus acquisition (Karl et al., 2008; Dyhrman et al., 2009; Beversdorf et al., 2010; Martínez et al., 2013; Carini et al., 2014; del Valle and Karl, 2014; Repeta et al., 2016; Horsman and Zechel, 2017; Sosa et al., 2017; Teikari et al., 2018).
In addition, it has been shown that marine snow particles are hotspots for MPn degradation and CH4 production (del Valle and Karl, 2014). MPn is produced abundantly by thaumarchaeotal ammonia-oxidizing archaea (AOA), cyanobacteria and some other marine microorganisms (Dyhrman et al., 2009; Metcalf et al., 2012; Van Mooy et al., 2015; Dang and Chen, 2017). In addition to bacteria, some archaea have also been found to harbor the gene encoding the carbon-phosphorus lyase, which is responsible for CH4 production from substrates such as MPn (Hove-Jensen et al., 2014).
The team work performed by different microorganisms during MPn production and consumption (Dang et al., 2013), particularly in phosphate-starved environments, may produce a large pool of surface seawater CH4 in the global ocean and contribute substantially to the net sea-to-air CH4 flux. Although CH4 production associated with MPn and particle microenvironments is intriguing and likely important, some alternative microbial processes have also been proposed to account for CH4 production in oxygenated seawater. Methane may also be produced during the microbial metabolization of dimethylsulfoniopropionate (DMSP) and its intermediate degradation products, such as dimethylsulfide (DMS), methanethiol (MeSH), and methylmercaptopropionate (MMPA) (Welsh, 2000; Damm et al., 2008, 2010; Florez-Leiva et al., 2013; Weller et al., 2013; Zindler et al., 2013).
Certain methanogens synthesize methylthiol:CoM methyltransferase or other substrate-specific CoM methyltransferases, using methylated sulfur compounds (MSCs, e.g., DMS, MeSH, and MMPA) as substrates for CH4 production (Tallant and Krzycki, 1997; Tallant et al., 2001; Fu and Metcalf, 2015). Many methanogens have been isolated and shown to grow with MSCs for methylotrophic methanogenesis (Kiene et al., 1986; Oremland et al., 1989; Finster et al., 1992; van der Maarel and Hansen, 1997; Lomans et al., 1999; Lyimo et al., 2000; Cha et al., 2013; Fu and Metcalf, 2015).
However, all these archaeal strains produce CH4 only under anoxic conditions. Whether and how the MSC-utilizing anaerobic methylotrophic methanogens can cope with oxygen stress under oxic conditions in the surface waters of the ocean is currently unresolved. A recent comparative genomic analysis revealed differences in oxygen tolerance among methanogenic archaea, suggesting that the Class II methanogens may be more adapted to microaerophilic environments than Class I methanogens (Lyu and Lu, 2018).
In addition, it has been proposed that certain aerobic bacteria may use DSMP as a carbon source and produce CH4 as a byproduct in oxic marine waters (Damm et al., 2010). The MSC-related CH4-producing mechanisms may constitute an important pathway in the ocean-to-atmosphere CH4 flux and in marine C and S cycling. DMSP is an abundant phytoplankton and coral metabolite that plays an important role in protecting organisms from osmotic, oxidative, and thermal stresses (Bullock et al., 2017).
Methane is a potent greenhouse gas (Bogard et al., 2014), while DMS is the most important biogenic climate-cooling gas in nature (Carini, 2016). Phytoplankton blooms are a global environmental and ecological problem in the coastal oceans of the world (Dang and Jiao, 2014; Dang and Lovell, 2016; Dang and Chen, 2017). With increasing anthropogenic impacts, such as excess nutrient discharges into the ocean, phytoplankton blooms will occur more frequently and extensively, particularly in estuarine and coastal seas (Jiao et al., 2014). More DMSP will therefore be produced by phytoplankton in the marginal seas in response to ocean eutrophication (Dang and Lovell, 2016).
Under this scenario, it is reasonable to hypothesize that the conversion of DMS to CH4 by marine microorganisms may further tip the balance of the Earth′s thermal budget and result in faster and stronger global warming (Florez-Leiva et al., 2013). In addition to the three major hypotheses regarding microbial processes and mechanisms for the paradoxical production of CH4 in oxic ocean waters mentioned above, some other hypotheses have also been proposed.
Terrestrial plants are a major source of CH4 (Conrad, 2009). Recently, the coccolithophore Emiliania huxleyi has been found to produce CH4 directly, without the involvement of archaeal methanogens or CH4-producing bacteria (Lenhart et al., 2016). Emiliania huxleyi is widespread and is the most abundant calcifying phytoplankton in the ocean where it plays an important role in ocean carbon sequestration (Krumhardt et al., 2017).
Its contribution to CH4 production in oxic surface waters, and thus to the ocean’s CH4 flux to the atmosphere requires an in-depth and systematic investigation. Damm et al. (2015) recently proposed another hypothesis about the mechanism of CH4 production in oxic nitrate-stressed marine environments. Bacterial respiration and low membrane permeability may create and maintain intracellular anaerobic conditions, which would be conducive to DMSP-dependent CH4 formation inside bacterial cells despite the oxic conditions of the surrounding bulk seawater (Damm et al., 2015).
This has been further supported by a modeling analysis of single-cell metabolic processes (Damm et al., 2015). Regarding the ocean methane paradox, it is likely that most of the microbial processes and mechanisms for CH4 production in oxic seawater proposed above may be valid. They may function under different environmental and/or biological conditions. However, it is still not quite clear what the environmental or biological determinants are that drive these differences.
Currently, their respective contributions to the seawater CH4 budget and sea-to-air CH4 emission are not clear, while their quantitative effect on the marine phosphorus and sulfur cycles also remains unclear. These uncertainties highlight the challenges that remain in marine microbiota and global change research and present opportunities for future scientific breakthroughs.
Future perspectives on marine CH4 research
Global climate change induced by greenhouse gases is a very complex process. There are complicated interactions and positive feedback effects involved that make global warming very difficult to mitigate. Methane as a potent greenhouse gas plays an important role in raising the global temperature. However, many features of the processes and mechanisms involved in the biogeochemical cycling relating to this simple organic molecule remain unresolved.
Methane emission via marine sediment gas hydrate dissociation constitutes a net source of atmospheric CH4. Recent studies have revealed certain unexpected ecological and climate impacts of CH4 released this way. It has recently been found that the upwelling produced by ascending CH4 gas bubbles moves deep-water nutrients into the surface water, and thus enhances photosynthetic CO2 fixation on the Svalbard margin (Pohlman et al., 2017). A recent study has indicated that massive emissions of CH4 from seep areas may be very common in shallow coastal seas (Borges et al., 2016).
Although CH4 emissions are unavoidably large, coastal marine environments, such as the Svalbard margin and others where similar CH4-related geological and ecological processes occur, may constitute net greenhouse gas sinks, and thus have a cooling effect on the regional and global climate (Pohlman et al., 2017). Several lessons have already been learned from history. One of these is that the real world is often more complex than was initially imagined.
There are many uncertainties and unknowns regarding marigenic CH4 in the process of global warming. Despite this, our understanding of the roles played by CH4 in ocean carbon cycling and global climate change has evolved. For example, regardless of what the processes and mechanisms are behind the paradoxical CH4 production in oxic marine waters, microbial activities always seem to be involved (Reeburgh, 2007). The diversity of microbes, their metabolic pathways, and environmental adaptation strategies results in complexity among the processes and functions of natural ecosystems, including CH4 production in oxic marine environments.
Under the influence of ongoing global and marine environmental changes, certain microbial processes may be altered. Global warming may have a direct impact on global microbial methanogenesis, which has been found to be more sensitive and responsive to a change in temperature than respiration and photosynthesis (Yvon-Durocher et al., 2014). With the projected rise in ocean temperature, it has been suggested that the increase in marine CH4 production and emission will be stronger than that of CO2 (Yvon-Durocher et al., 2014). Increased anthropogenic activities and ocean warming will exacerbate the hypoxic conditions in most estuarine and coastal seas and in oceanic oxygen minimum zones (Gruber, 2011; Gilly et al., 2013; Dang and Jiao, 2014; Dang and Lovell, 2016; Dang and Chen, 2017).
Some of the hypoxic waters may develop anoxic cores that will facilitate microbial CH4 production. Under the influence of global climate change, upwelling in certain areas of the ocean will be enhanced (Sydeman et al., 2014; Wang et al., 2015). This may create hypoxic and even anoxic water bodies that facilitate methanogenesis (Bakun, 2017). Strengthened upwelling also stimulates the formation of phytoplankton blooms (Kudela et al., 2010), enhancing CH4 production via the enhanced production of DMSP and increased formation of marine particles (Damm et al., 2008, 2010; Florez-Leiva et al., 2013; Weller et al., 2013; Dang and Lovell, 2016).
Moreover, increased ocean acidification may reduce nitrification in the ocean, because more environmental ammonia would be converted to ammonium, which would reduce the ammonia oxidation rate by AOA and ammoniaoxidizing bacteria (AOB) in both seawater and sediments (Beman et al., 2011; Braeckman et al., 2014; Dang and Chen, 2017). How this may influence the production of MPn by AOA (but not by AOB) and the related production of CH4 is currently unknown. Furthermore, methanogenic and methanotrophic microbes control the production and consumption of CH4, and thus its abundance and dynamics in marine environments. Certain trace elements (e.g., Co, Cu, Fe, and Ni) are important cofactors for enzymes involved in microbial methanogenesis and methanotrophy (Glass and Orphan, 2012; DiSpirito et al., 2016; Paulo et al., 2017; Semrau et al., 2018).
The bioavailability of these trace elements in the ocean may play an important role in the microbial activities involved in CH4 production and consumption. On the other hand, heavy metals may exert negative impacts on the activities of microbial methanogens and methanotrophs via enzyme toxification. Ocean warming, acidification, deoxygenation, eutrophication, and pollution may all influence the availability of trace elements, and the effects of heavy metals in marine environments (Dang and Chen, 2017).
The association of microbial activity with marine inorganic elements and contaminants makes the prediction of changes in the future climate and marine environment even more complex and difficult. The advancement of “omics” techniques, such as metagenomics, metatranscriptomics, and metaproteomics, provides opportunities for the study and understanding of the microbes and their processes and mechanisms in the ocean’s CH4 dynamics (Lloyd, 2015). The combination of the “omics” analyses with in situ flux measurements may enable the development of more powerful and accurate biogeochemical models, resulting in a better prediction of the ocean’s CH4 behavior and its influence on global change.
EARTH CLIMATE covers the broad spectrum of climate change, and the solutions, with the focus on the sciences. Earth Climate – we endorse data, facts, empirical evidence.
We create visually compelling, accessible science content that transforms complex research into engaging videos and articles, helping millions understand the discoveries shaping our world. [Paypal | Patreon]


{kind=link}
{kind=link}
{kind=link}
